Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность
процесса, значениеСтраница 1

В анаэробных условиях пировиноградная кислота (пируват) подвергается дальнейшим превращениям в ходе спиртового, молочнокислого и других видов брожений, при этом NADH используется для восстановления конечных продуктов брожения, регенерируя в окисленную форму. Последнее обстоятельство поддерживает процесс гликолиза, для которого необходим окисленный NAD + . В присутствии достаточного количества кислорода пируват полностью окисляется до С02 и Н20 в дыхательном цикле, получившем название цикла Кребса или цикла ди- и трикарбоновых кислот. Все участки этого процесса локализованы в матриксе или во внутренней мембране митохондрий.
Последовательность
реакций в цикле Кребса.Участие органических кислот в дыхании давно привлекало внимание исследователей. Еще в 1910 г. шведский химик Т. Тунберг показал, что в животных тканях содержатся ферменты, способные отнимать водород от некоторых органических кислот (янтарной, яблочной, лимонной). В 1935 г. А. Сент-Дьердьи в Венгрии установил, что добавление к измельченной мышечной ткани небольших количеств янтарной, фумаровой, яблочной или щавелевоуксуснсй кислот резко активирует поглощение тканью кислорода.
Учитывая данные Тунберга и Сент-Дьердьи и исходя из собственных экспериментов по изучению взаимопревращения различных органических кислот и их влияния на дыхание летательной мышцы голубя, английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до С02 через «цикл лимонной кислоты» да счет отнятия водорода. Этот цикл и был назван его именем.
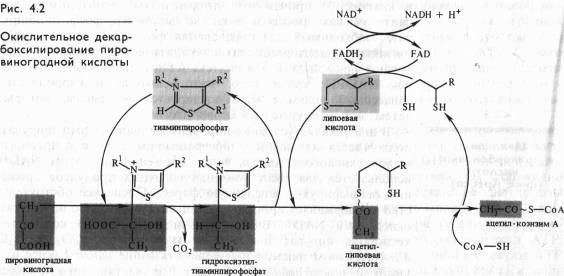
Непосредственно в цикле окисляется ле сам пируват, а его производное — ацетил-СоА. Таким образом, первым этапом на пути окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мультифёрментного комплекса. В состав его входят три фермента и пять коферментов. Коферментами служат тиаминпирофосфат (ТПФ) — фосфорилированное производное витамина Вь липоевая кислота, коэнзим A, FAD и NAD+. Пируват взаимодействует с ТПФ (декарбоксилазой), при этом отщепляется С02 и образуется гидроксиэтильное производное ТПФ (рис. 4.2). Последнее вступает в реакцию с окисленной формой липоевой кислоты. Дисульфидная связь липоевой кислоты разрывается и происходит окислительно-восстановительная реакция: гидроксиэтильная группа, присоединенная к одному атому серы, окисляется в ацетильную (при этом возникает высокоэнергетическая тиоэфирная связь), а другой атом серы липоевой кислоты восстанавливается. Образовавшаяся ацетиллипоевая кислота взаимодействует с коэнзимом А, возникают ацетил- СоА и восстановленная форма липоевой кислоты. Водород липоевой кислоты переносится затем на FAD и далее на NAD + . В результате окислительного декарбоксилирования пирувата образуются ацетил-СоА, С02 и NADH.
Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса. Цикл Кребса начинается с взаимодействия ацетил-СоА с енольной формой щавелевоуксусной кислоты. В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота. Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой. В первой реакции в результате дегидратации лимонной кислоты образуется цис-аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием NAD- или NADP-зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием а-кетоглутаровой кислоты (а-оксоглутаровой кислоты).
а-Кетоглутарат, подобно пирувату, подвергается реакции окислительного декарбоксилирования. а-Кетоглутаратдегидрогеназный мультиэнзимный комплекс сходен с рассмотренным выше пируватдегидрогеназным комплексом. В ходе реакции окислительного декарбоксилирования а-кетоглутарата выделяется С02, образуются NADH и сукцинил-СоА.
Подобно ацетил-СоА, сукцинил-СоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-СоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-CoA может трансформироватся в образование фосфатной связи АТР. При участии сукцинил- СоА-синтетазы из сукцинил-СоА, ADP и Н3Р04 образуются янтарная кислота (сукцинат), АТР, регенерирует молекула СоА. АТР образуется в результате субстратного фосфорилирования.
Это интересно:
Ошибочные «признаки» ядовитых грибов
Народные приметы, «позволяющие определить ядовитые грибы», основаны на разнообразных заблуждениях и не позволяют судить об опасности грибов:
1.Ядовитые грибы имеют неприятный запах, а съедобные — приятный (запах бледной поганки почти иде ...
Цитопатогенное действие
Характерное для РСВ цитопатогенное действие - это образование синцития. В синцитиях ядра клеток часто образуют агрегаты, окружающие центральную полость. Однако формирование синцития наблюдается далеко не всегда; характер ЦПД определяется ...
Признаки биологического возраста
Не любой признак, изменяющийся с возрастом, может определять биологический возраст человека. В случае старения кожи, появления седины и морщин, функционирование остальных органов, особенно мозга и сердца остается на высоком уровне, тогда ...